Melatonina: azioni ed utilità nell'uomo
di Maurizio Nordio
Dipartimento di Fisiopatologia Medica, Università “Sapienza”, Roma
La vita sulla Terra, fin dal suo inizio, è stata sempre regolata dalla successione ritmica del giorno e della notte, oltre che delle stagioni, dipendente dalla presenza della radiazione luminosa del sole. Pertanto non stupisce il fatto che gli organismi viventi abbiano sviluppato sistemi assai efficaci in grado di percepire la ciclicità dell’ambiente esterno e quindi la presenza (o assenza) della luce e la sua durata nelle 24 ore e adattare di conseguenza le proprie attività giornaliere e stagionali alle variazioni ritmiche dell’ambiente circostante. Tale capacità di adattamento è, in ultima analisi, la chiave di volta per una migliore probabilità di sopravvivenza e quindi per la massima diffusione della specie. Quindi, in termini di efficacia del processo evolutivo, l’organismo in grado di adattarsi meglio ha maggiori probabilità di resistere con successo alle mutevoli condizioni ambientali. La corretta espressione di tale capacità necessita innanzitutto della presenza di un sistema endogeno che, come detto, possa riconoscere le variazioni cicliche dell’ambiente circostante e comunicarle alle strutture incaricate della risposta. In questo senso, sia nell’animale che nell’uomo, il cammino dell’evoluzione ha determinato lo sviluppo di un meccanismo molto fine in grado di registrare appunto le modificazioni ritmiche dell’ambiente, soprattutto in termini di alternanza luce/buio, farle giungere facilmente ai centri deputati alla decodifica e stimolare una risposta tale per cui il messaggio possa arrivare in periferia alla singola cellula e determinare una risposta in senso adattativo che sia in sincronia con le caratteristiche dell’ambiente esterno.
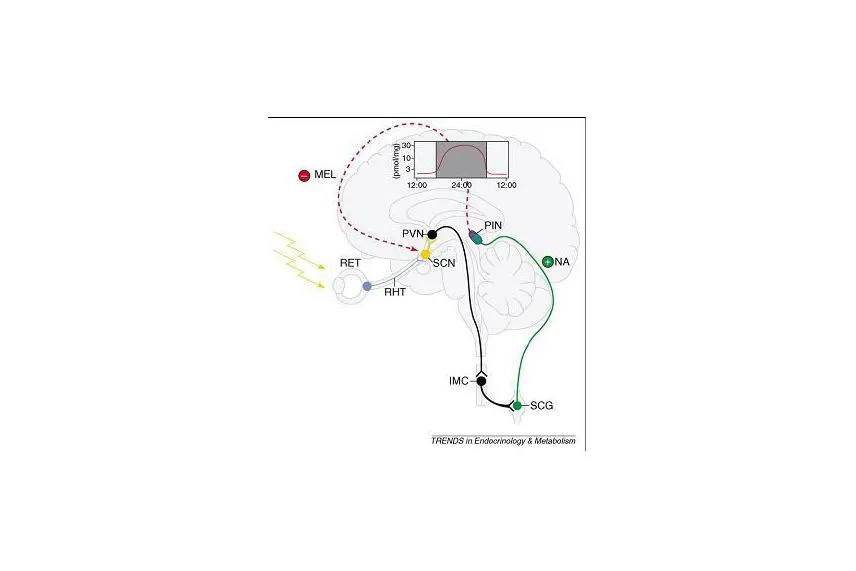
In tal senso, è ben noto che la luce viene considerata il più potente sincronizzatore dei ritmi circadiani dell’uomo (Middleton et al. 2002). Essa trasporta 2 tipi di informazioni differenti, entrambe importanti per la sopravvivenza dell’organismo considerato: la prima riguarda informazioni relative alla forma tridimensionale degli oggetti, al colore e alla percezione del movimento; la seconda è costituita da un segnale temporale riguardante la durata del giorno e della notte. A causa di ciò, durante il processo evolutivo si sono sviluppati 2 diversi sistemi di visione in tutti i vertebrati, in grado di distinguere i 2 tipi di informazioni trasportate attraverso la radiazione luminosa: il cosiddetto “occhio laterale” (che viene usato per la formazione delle immagini degli oggetti ed il cosiddetto “terzo occhio”, in grado di rilevare lo stimolo luminoso in termini di alternanza luce/buio. Negli animali più evoluti e nell’uomo tale ultima struttura viene identificata con la ghiandola pineale. Inoltre, studi molto recenti hanno potuto documentare la presenza, sulla superficie della retina, di una popolazione di cellule non adatte alla visione, diverse dai “coni” e “bastoncelli”, ma in grado di reagire allo stimolo luminoso e le cui fibre, attraverso il tratto retinoipotalamico, raggiungono il nucleo soprachiasmatico (Gehering, 2005) il quale viene ormai considerato il componente principale del cosiddetto “orologio biologico” dell’organismo, con il compito di generare i ritmi circadiani tramite le istruzioni presenti nel genoma (Buijs et al., 2003) in grado di attivare ritmicamente una serie assai complessa di geni con il compito di codificare le proteine utili per la trasmissione del segnale che determina l’attivazione dei meccanismi intracellulari.
Tuttavia, possedere una propria ritmicità circadiana intrinseca (che nell’uomo è di circa 24.2 ore) non è sufficiente per il corretto funzionamento dell’organismo; infatti è importante che tale ciclicità sia in sincronia con i ritmi dell’ambiente circostante. A questo scopo, il messaggio ritmico generato dal nucleo soprachiasmatico viene successivamente trasmesso ad una struttura endocrina in grado di riceverlo e reagire allo stimolo nervoso mediante la produzione ormonale che viene diffusa a livello sistemico. Si tratta della ghiandola pineale (o epifisi) e del suo principale prodotto di secrezione, la melatonina che chimicamente è un indolo (N-acetil-5-metossitriptamina) e deve il proprio nome agli effetti sulla pigmentazione cutanea degli anfibi. L’interconnessione di strutture quali la retina, il nucleo soprachiasmatico e la pineale costituisce ciò che viene comunemente definito come “sistema fotoneuroendocrino” (Korf et al., 1998)(Fig.1).
LA GHIANDOLA PINEALE
Unico organo endocrino in contatto con l’ambiente esterno tramite una complessa via nervosa precedentemente ricordata (e per questo appunto denominato anche trasduttore fotoneuroendocrino), l’epifisi svolge un’attività apparentemente assai semplice e cioè quella di produrre sostanze, fra le quali la più importante è la melatonina, in risposta a stimoli provenienti dal nucleo soprachiasmatico. In realtà esiste una sorta di discrepanza fra tale “semplice” attività della pineale e la moltitudine di geni espressi ritmicamente e di segnali intracellulari coinvolti in tale meccanismo. Dati recentissimi della letteratura scientifica suggeriscono che questa apparente discrepanza non sia altro se non, da un lato la conferma dell’importanza che riveste la capacità di mantenere una adeguata ritmicità e sincronia delle funzioni endogene per il buon funzionamento dell’organismo, dall’altro il preludio alla identificazione di attività della pineale finora ignote (Maronde e Stehle, 2007).
La produzione di melatonina nel circolo periferico avviene seguendo un ben noto ritmo circadiano che prevede valori plasmatici massimi (dell’ordine di circa 50-200 pg/ml) durante le ore notturne e minimi durante quelle diurne (circa 2-10 pg/ml). Le fibre post-gangliari del sistema simpatico provenienti dal ganglio cervicale superiore (che è collegato con la retina) terminano a livello della membrana del pinealocita e regolano la sintesi di melatonina mediante la liberazione di noradrenalina (o norepinefrina: NE) dal bottone sinaptico. La liberazione di NE avviene durante le ore notturne. Successivamente al legame della NE con il recettore ?-adrenergico sulla membrana del pinealocita si ha l’attivazione dell’enzima adenilato-ciclasi e conseguente produzione di AMP-ciclico. Tale aumento promuove la sintesi di proteine, fra le quali anche gli enzimi necessari per la sintesi di melatonina e in particolare l’enzima limitante N-acetiltransferasi (AA-NAT)(Klein, 2004). Durante la fase di luce, l’attività elettrica del nucleo soprachiasmatico è elevata e in tali condizioni la liberazione di NE risulta bassa. Al contrario, durante la fase di buio l’attività del SCN viene inibita, mentre la sintesi della melatonina è stimolata dall’aumento della NE. All’interno del pinealocita, in seguito a stimolazione dei recettori adrenergici sulla membrana cellulare, a partire dal triptofano, si ha la formazione di composti intermedi, fra i quali anche la serotonina, in presenza di una serie di enzimi, di cui il principale è l’AA-NAT che converte la serotonina in N-acetilserotonina, la cui attivazione ciclica è direttamente dipendente dall’attività del nucleo soprachiasmatico, dalla informazione luminosa, oltre che dalla ritmica trascrizione dei geni codificanti (Klein, 2007). Ne consegue che anche i soggetti non vedenti, pur non percependo lo stimolo luminoso attraverso la retina, e quindi mancando del segnale più efficace in grado di inibire la secrezione di melatonina, dimostrano ugualmente un ritmo circadiano (e non una secrezione di melatonina sempre elevata come erroneamente si potrebbe pensare), anche se spesso tale ritmo non è del tutto sincronizzato con l’ambiente esterno (Bellastella et al., 1998). Una volta formata, la melatonina non viene immagazzinata all’interno della ghiandola pineale, ma diffonde nei capillari e nel fluido cerebrospinale.
E’ interessante notare come ciascun individuo adulto abbia livelli massimi di melatonina caratteristici della propria persona e molto stabili nel tempo in termini di concentrazione plasmatica, e come essi possano essere anche molto diversi da quelli degli altri soggetti (Arendt, 1988). Conseguentemente, sarà irrilevante sapere se un determinato individuo presenti un picco notturno di melatonina di 100 pg/ml piuttosto che di 200 pg/ml. Al contrario, acquisiscono grande valore sia la presenza (o l’assenza) di un ritmo circadiano ben evidente, sia la durata del periodo in cui la melatonina si mantiene più elevata rispetto ai valori diurni, sia le caratteristiche di anticipo o di ritardo della secrezione di melatonina rispetto alle fasi della giornata (Reiter, 1989). La melatonina raggiunge facilmente tutti i tessuti dell’organismo in tempi assai brevi. La curva di dimezzamento nel sangue ha un aspetto bi-modale, con un primo picco di distribuzione dopo 2 minuti ed un secondo dopo circa 20 minuti (Claustrat et al., 2005). La melatonina circolante viene metabolizzata prevalentemente dal fegato, dove viene dapprima idrossilata dagli enzimi della famiglia del citocromo P-450 (CYPIA2 e CYPIA1) e successivamente coniugata con solfato per essere poi escreta con le urine sottoforma di 6-sulfossimelatonina. Altrettanto importante è però il metabolismo extra epatico della melatonina. In particolare, tale indolo può essere metabolizzato anche in maniera non-enzimatica in tutte le cellule ed anche nel compartimento extracellulare. Esempi di ciò sono il tessuto cerebrale, dove la melatonina viene convertita in derivati chinuraminici (il più importante è l’N1-acetil-N2-formil-5-metossichinuramina: AFMK) che conservano alcune proprietà della molecola di partenza, quali l’attività antinfiammatoria e antiossidante, e i radicali liberi, in grado di convertire la melatonina in 3-idrossimelatonina.
IL RUOLO DELLA LUCE
La presenza di un sistema di sincronizzazione così sofisticato e potente suggerisce l’importanza che riveste, per l’organismo umano e non, il mantenere una serie di funzioni ritmiche in sincronia con l’ambiente che ci circonda (Liu et al., 2000). Ad ulteriore conferma di ciò, del tutto recentemente è stata documentata, nell’animale da esperimento, la presenza di un sistema in grado di fornire all’organismo informazioni specifiche riguardanti la cosiddetta ritmicità circannuale (in pratica la possibilità di riconoscere l’alternanza delle stagioni). Il corretto funzionamento di tale sistema sembra dipendere dall’interazione fra le cellule del pace-maker endogeno, sincronizzate dalla melatonina e le cellule ipofisarie prolattino-secernenti (Lincoln et al., 2006). Inoltre, è necessario sottolineare che anche in assenza dello stimolo luminoso la ritmicità endogena, generata a livello del nucleo soprachiasmatico e comunicata alla pineale, viene conservata ancorché non perfettamente sincronizzata con l’esterno. Ne consegue ad esempio che anche i soggetti non vedenti, pur non percependo lo stimolo luminoso e quindi mancando del segnale più efficace in grado di inibire la secrezione di melatonina, dimostrano ugualmente un ritmo circadiano, anche se spesso non del tutto sincronizzato con l’ambiente esterno (Bellastella et al., 1998).
A supporto della stretta correlazione fra il ritmo luce/buio e la secrezione ciclica della melatonina, esiste tutta una serie di evidenze sperimentali indicanti che l'esposizione ad una fonte luminosa di intensità e lunghezza d'onda adeguate, durante il periodo notturno, determina una drastica e reversibile riduzione dei livelli di melatonina fino ai valori diurni. In tal senso, i dati della letteratura indicano come più efficace una lunghezza d’onda compresa fra i 420 e i 480nm (luce blu e verde), mentre meno efficace risulta essere la lunghezza d’onda nell’ambito del giallo e del rosso. Inoltre, la maggior parte degli sono concordi nel ritenere che si debba superare un valore-soglia (circa 300-500 lux, nell’uomo)(Reiter, 1989; Skene e Arendt, 2006), anche se dati recentissimi indicano che è possibile modificare il ritmo circadiano sottoponendo l’animale da esperimento ad una intensità luminosa estremamente bassa (di poco superiore alla luce della luna)(Evans et al., 2007). Tuttavia altri autori hanno dimostrato che la riduzione della secrezione di melatonina dipendente non soltanto dal livello di intensità della radiazione luminosa, ma anche dal tempo di esposizione. In particolare è stato possibile documentare come l’esposizione ad una intensità luminosa di soli 200 lux fosse in grado di ridurre progressivamente le concentrazioni plasmatiche di melatonina, con l’aumentare del tempo di esposizione alla luce. Malgrado ciò, l’inibizione della secrezione di melatonina indotta dalla esposizione alla luce è comunque dose-dipendente, con le intensità maggiori in grado di determinare soppressione maggiore (Lack e Wright, 2007).
Infine, recentemente è stato dimostrato come, ai fini della capacità melatonino-soppressiva della luce, abbia importanza anche la cosiddetta “storia luminosa” del soggetto e quindi che tale sistema sia in grado di adattarsi. Ciò ha grande importanza nella società odierna che è caratterizzata da quello che viene comunemente definito come “inquinamento luminoso” e cioè la costante esposizione degli individui a radiazione luminosa durante le ore notturne. Dati della letteratura sempre più numerosi indicano infatti un collegamento fra esposizione alla luce durante le ore notturne, riduzione dei livelli di melatonina e rischio di malattia, come riportato più avanti (ad esempio l’aumento dei casi di tumore mammario nelle donne “turniste”)(Schernhammer et al., 2006, a). Tale proprietà soppressiva della luce viene comunemente sfruttata a scopo terapeutico (fototerapia) in alcune situazioni di desincronizzazione, quali la sindrome da spostamento del fuso orario (jet-lag)(Waterhouse et al., 1997) o nei soggetti sottoposti a turni di lavoro durante le ore notturne (turnisti)(Arendt et al., 1997, a) ed infine in alcune patologie di pertinenza psichiatrica come la depressione stagionale e la bulimia nervosa (Partonen e Lönnqvist, 1998). Inoltre, dal momento che la luce è una radiazione elettromagnetica, è stato ipotizzato che anche l’esposizione a campi elettromagnetici (ELF) potesse modificare la secrezione di melatonina. Studi sperimentali nell’animale e nell’uomo hanno dimostrato la validità di tale postulato (Ronco e Halberg, 1996; Burch et al., 1998). Più in particolare, nostri dati recenti, ottenuti tramite l’impiego di un protocollo di lavoro standardizzato per lo studio degli effetti biologici dei campi elettromagnetici (Benedetti et al., 2005) in due gruppi di soggetti adulti (esposti e non esposti a ELF presso la loro residenza), hanno dimostrato la correlazione inversa fra presenza di sintomatologia soggettiva in caso di esposizione a ELF e concentrazioni urinarie di 6-sulfossimelatonina (dati non pubblicati).
MODALITA’ DI TRASFERIMENTO DEL SEGNALE “MELATONINA”
L’azione della melatonina a livello periferico può essere svolta tramite una serie di recettori in grado di venire attivati dall’indolo pinealico e localizzati sulla membrana (MT1 e MT2) e sul nucleo delle cellule epifisarie (Steinhilber et al., 1995; Vanecek, 1998). I primi due appartengono alla famiglia dei recettori accoppiati alle proteine “G” e sono responsabili degli effetti a livello del nucleo soprachiasmatico. In particolare, l’MT1 agisce sopprimendo l’attività elettrica di tale nucleo ipotalamico, mentre l’MT2 agisce inducendo variazioni della fase del ritmo circadiano. Entrambi i recettori sono espressi anche a livello periferico, dove svolgono molteplici azioni, quali ad esempio quelle immunologiche o di controllo vasomotorio (MT1: vasocostrizione; MT2: vasodilatazione) (Dubocovich e Markowska, 2005). La melatonina si lega anche ai recettori nucleari appartenenti alla famiglia dei recettori ad acido retinoico (RORα1, RORα2 e RZRβ) i quali sono coinvolti nel meccanismo di immunomodulazione (RORα1 e RORα2) e vengono espressi anche a livello del tessuto cerebrale (RZRβ)(Pandi-Perumal et al., 2006). Allo scopo di chiarire meglio il meccanismo d’azione della melatonina, è importante sottolineare che anche la sensibilità dei recettori per l’indolo dimostra una chiara variazione circadiana, identificando nel tardo pomeriggio, cioè subito prima dell’inizio del fisiologico incremento notturno della melatonina, il momento di maggiore sensibilità in tal senso. Al contrario, durante il periodo diurno si assiste ad una marcata riduzione di detta sensibilità, determinata dalla massiccia presenza dell’indolo durante la notte precedente (fenomeno comune a tutti i recettori per gli ormoni e noto come down-regulation). Pertanto, un’eventuale assunzione di melatonina nelle prime ore del giorno potrebbe risultare meno efficace a causa della impossibilità, per tale sostanza, di legarsi in quantità sufficiente ai recettori e quindi di svolgere i propri effetti biologici (Hazlerigg et al., 1993; Gerdin et al., 2004).
Tuttavia, le azioni dell’indolo pinealico a livello cellulare non dipendono soltanto dalla presenza dei recettori periferici per la melatonina, ma anche dalla capacità che tale sostanza ha di diffondere rapidamente all’interno dei tessuti e raggiungere virtualmente ogni singola cellula dell’organismo, senza la necessità di un legame recettoriale, grazie alla semplicità della molecola di melatonina, associata alle sue piccole dimensioni (basso peso molecolare) ed alla sua liposolubilità. Ad ulteriore sostegno dell’importanza della melatonina prodotta da strutture diverse dalla pineale, certamente maggiore di quanto sostenuto fino ad oggi, da un lato la non completa scomparsa della melatonina dal sangue in seguito ad ablazione chirurgica della pineale, dall’altro il rilievo abbastanza recente della identificazione di siti di produzione della melatonina diversi dall’epifisi e del tutto autonomi, quali ad esempio la retina, la ghiandola di Harder (presente solo in alcune specie animali, non nell’uomo), i megacariociti, le piastrine (Huether, 1992), l’epitelio gastrointestinale. In quest’ultimo caso, è ormai ben noto che la melatonina non solo è presente in tutte le cellule del tratto gastrointestinale, ma viene attivamente prodotta dalle cellule enterocromaffini, raggiungendo concentrazioni maggiori di quelle della pineale stessa e del sangue periferico. Inoltre può essere liberata nel circolo periferico e la sua produzione risulta aumentata in special modo in risposta alla introduzione di cibo (Bubenik, 2002). A livello intestinale è in grado di incrementare la secrezione di bicarbonati dalla mucosa duodenale, attraverso un’azione mediata dai recettori MT2, allo scopo di proteggere la mucosa duodenale dalla secrezione acida gastrica. A conferma di tale meccanismo, dati recenti hanno dimostrato una correlazione inversa fra concentrazioni urinarie di 6-sulfossimelatonina e incidenza di recidive di ulcere duodenali nell’uomo (Malinovskaya et al., 2001). Inoltre la melatonina è stata identificata anche nella bile, a concentrazione anche di circa 1000 volte maggiori rispetto a quelle plasmatiche diurne. E’ stato ipotizzato che tali azioni della melatonina a livello del tratto gastrointestinale siano necessarie per fornire una protezione antiossidante all’epitelio gastrointestinale e contribuire a rafforzare il meccanismo di protezione delle cellule epiteliali dall’aggressione dell’acido cloridrico e degli acidi biliari.
AZIONI PERIFERICHE DELLA MELATONINA
Come già ricordato, la melatonina è una molecola in grado di svolgere molteplici azioni fisiologiche, segnalando non soltanto il momento del giorno o la stagione dell’anno, ma anche dimostrando evidenti proprietà di immunomodulazione, di citoprotezione e altro. L’importanza di questa molecola è tale de essere considerata come uno dei primi segnali biologici che siano apparsi sulla Terra (Claustrat et al., 2005). Segnale biologico che svolge, in particolare, un ruolo di primaria importanza nel meccanismo che: regola i ritmi dell’organismo e li mantiene in sincronia con quelli dell’ambiente esterno; svolge potente attività di protezione cellulare contro gli eventi ossidativi; dimostra attività di stimolazione del sistema immunitario; ha effetti riequilibranti sulla funzione cardiovascolare, quella di rimaneggiamento dell’osso, sulla regolazione della massa corporea e sul sistema riproduttivo.
Melatonina e ritmo sonno/veglia
Dal momento che la melatonina viene prodotta di notte, la sua azione è stata spesso associata al sonno. Tuttavia, è ormai ben chiaro che tale indolo non può essere considerato un induttore universale del sonno. Infatti esistono animali cosiddetti notturni e che svolgono quindi la loro attività essenzialmente di notte, i quali dimostrano livelli plasmatici di melatonina elevati durante quel periodo. Più correttamente, la melatonina è un composto ubiquitario correlato alle ore notturne, in grado di segnalare la durata del periodo di buio. E’ pertanto logico ritenere che fenomeni comportamentali in senso lato quali il sonno, la riduzione della temperatura corporea, del grado di attenzione, della performance e la modificazione delle funzioni metaboliche durante le ore notturne siano connessi e influenzati dalla presenza della melatonina. A conferma di ciò, è ormai ben noto che le curve della temperatura, del grado di attenzione e di performance sono inversamente correlate a quella della melatonina. Inoltre, la somministrazione dell'indolo pinealico a volontari sani nelle ore diurne, ha permesso di documentare una riduzione della temperatura corporea dopo circa 2 ore dalla somministrazione (Stone et al., 2000). Allo stesso modo, la somministrazione di un’adeguata quantità di luce a soggetti normali durante il periodo notturno determina un incremento della temperatura corporea e del livello di attenzione (Badia et al., 1991). Per quanto riguarda i soggetti anziani, dati recenti indicano che la somministrazione di melatonina è in grado di migliorare la qualità del ritmo della temperatura corporea, rendendolo simile a quello dei soggetti giovani (Gubin et al., 2006). La relazione temporale fra l’aumento notturno dei livelli di melatonina endogena e l’ingresso nelle prime fasi del sonno consente di ipotizzare che tale indolo faciliti il sonno inibendo il cosiddetto meccanismo circadiano della veglia tramite un effetto medianto dai recettori MT1 presenti nel nucleo soprachiasmatico (Lavie, 1997).
Per quanto riguarda il ritmo sonno/veglia, sono ormai innumerevoli i dati della letteratura scientifica indicanti la capacità della melatonina di modificare temporalmente le fasi del sonno, avanzandole o ritardandole in dipendenza del momento di somministrazione esogena. Tale capacità, attribuita all’azione dei recettori MT2 nell’SCN, viene normalmente utilizzata in terapia in tutte quelle condizioni in cui il ritmo sonno/veglia non risulti più sincronizzato con il ritmo luce/buio dell’ambiente in cui ci si trova (Bergstrom e Hakanson, 1998; Cassone, 1998). Evidenze sperimentali indicano, ad esempio, che in particolari condizioni (variazione rapida del fuso orario, turni di lavoro notturni, invecchiamento, sindrome da ritardo, o avanzamento, della fase del sonno, depressione stagionale, ecc.) si possa assistere, appunto, ad una desincronizzazione del sonno e delle altre funzioni ritmiche rispetto all'ambiente esterno, con perdita dello stato di benessere (Lewy e Sack, 1997). In pratica, il tentativo di addormentarsi in un momento del ciclo circadiano che non è ottimale per l’attivazione di un episodio di sonno, determina di addormentamento o risveglio precoce. In ultima analisi ciò determina una riduzione del tempo totale di sonno, causando di fatto una perdita di benessere generale. Nel momento in cui ciò si verifica troppo velocemente nella persona sana (come nel caso di uno spostamento del fuso orario in occasione di voli intercontinentali), si può manifestare la cosiddetta “sindrome da jet-lag” che, in ultima analisi, rende evidente la difficoltà del nostro organismo a “comprendere” le caratteristiche di ritmicità del nuovo ambiente, determinando la comparsa di una serie di sintomi più o meno gravi, variabili dal semplice malessere, fino all’insonnia in un elevato numero di soggetti (fino al 94% di essi)(Harma et al., 1994; Lack e Wright, 2007). Gli effetti negativi maggiori si verificano durante i voli verso est perché ciò richiede un avanzamento della fase dell’orologio biologico, anziché un ritardo. Il ruolo della melatonina nella "sindrome da jet-lag" è ormai universalmente accettato, anche perché la somministrazione di tale indolo, secondo precisi protocolli, è in grado di annullare i disturbi (compresi quelli del sonno) dipendenti dalle variazioni del fuso orario (Arendt et al., 1997). In particolare, induce l’avanzamento della fase di circa 1.1-1.4 ore ogni giorno di somministrazione; pertanto, la completa risincronizzazione (spostamento della fase di circa 7-8 ore) avviene in media dopo 5 giorni di assunzione, riducendo nel contempo, anche i sintomi correlati (Arendt, 2005). Infine, dati recenti indicano che anche l’esposizione alla luce, somministrata nel momento opposto rispetto alla melatonina, è in grado di contribuire alla risincronizzazione delle fasi del sonno (Cardinali et al., 2006).
Accanto a tali informazioni, i dati della letteratura indicano un ruolo per l'indolo pinealico anche nei disturbi del ritmo sonno/veglia dipendenti da altre cause. Più in particolare, è stato possibile documentare come la riduzione delle concentrazioni massime notturne di melatonina possa determinare un'alterazione del ritmo del sonno nell'uomo (Haimov et al., 1994). A tale proposito è necessario ricordare che le concentrazioni massime notturne di melatonina nel sangue periferico dei soggetti normali non sono costanti per tutta la vita. Esse si riducono progressivamente con l’età ed infatti il bambino, fino al momento della pubertà, dimostra i livelli notturni più elevati. Successivamente, durante il periodo puberale la melatonina si riduce per raggiungere i valori dell’adulto i quali si mantengono tali fino all’età di circa 50-55 anni, quando iniziano a ridursi ulteriormente e progressivamente, per giungere anche alla scomparsa della ritmicità circadiana in età molto avanzata (Touitou, 1995; Zhou et al., 2003). In tale ultimo gruppo è possibile documentare frequentemente un’alterazione dell’architettura, della qualità e della durata del sonno che viene annullata dalla somministrazione di melatonina (Hughes et al., 1998). In accordo con tali considerazioni, è stato dimostrato come la riduzione delle concentrazioni plasmatiche notturne di melatonina sembri significativamente più pronunciata nelle donne durante il periodo menopausale (Vakkuri et al., 1996). Ulteriori dati recenti indicano che in un gruppo di soggetti in menopausa, il ritmo circadiano della melatonina risulta avanzato di circa 30 minuti rispetto ai soggetti di età inferiore. In tal senso, i soggetti osservati nello studio riferivano la necessità di riposo, seguito da addormentamento, nelle prime ore serali e risveglio precoce (segni di sfasamento del ritmo circadiano)(Yoon et al., 2004). Inoltre, la valutazione della distribuzione oraria delle vampate nei soggetti in menopausa, ha permesso di documentare come tale frequenza sia mediamente più elevata durante le ore pomeridiane, periodo della giornata durante il quale classicamente i livelli plasmatici di melatonina raggiungono il minimo (Freedman et al., 1995). Concettualmente simile il discorso riguardante le donne con sindrome premestruale (PMS= Pre-Menstrual Syndrome), in quanto dati della letteratura hanno potuto documentare come in tali soggetti esista una associazione fra sintomi riferiti (alterazioni dell’umore, modificazioni del ritmo del sonno, ecc.) e alterazioni del ritmo di melatonina, sia in termini di sfasamento di esso, sia in termini di riduzione della secrezione globale giornaliera, significativamente più evidente durante la fase luteale, rispetto a quella follicolare (Parry et al., 1997; Barron 2007).
Melatonina e protezione contro gli eventi ossidativi
Rappresentanti di una categoria di sostanze, forse le più dannose per le cellule, sono i cosiddetti "radicali liberi". Anche se il termine è errato per definire tutta la categoria, perchè appartiene soltanto ad una ristretta e ben specifica serie di elementi, viene ormai generalmente individuato come caratteristico di composti nocivi in generale, derivati dal metabolismo dell’ossigeno che si formano sia come conseguenza delle normali funzioni metaboliche dell’organismo, sia derivanti dall’influenza di fattori esterni sull’organismo (raggi ultravioletti, raggi X, inquinamento ambientale, additivi alimentari, fumo di sigaretta, ecc.). La loro pericolosità deriva essenzialmente dal fatto che sono molto instabili chimicamente e quindi reagiscono rapidamente con qualunque molecola si trovi nelle vicinanze. Tale fatto, che tecnicamente consiste nel "furto" di un elettrone da parte del radicale libero nei confronti di una qualsiasi altro composto, determina il danneggiamento di grado variabile, da lieve fino alla completa distruzione, di molecole di ogni tipo (lipidi, proteine, acidi nucleici, ecc.). Il danno risulta tanto più grave quanto più importante è la molecola danneggiata. In particolare, la produzione di radicali liberi attiva una serie di meccanismi che agiscono per determinare un insulto cellulare più o meno esteso e che comprende: a) la reazione con gli acidi nucleici, i nucleotidi, i polisaccaridi e le proteine; b) il legame covalente con i componenti la membrana cellulare (lipidi, proteine, enzimi, recettori e sistemi di trasporto); c) l'inizio della perossidazione lipidica; d) l'alterazione del citoscheletro; e) la morte cellulare.
Tuttavia un danno irreversibile non si manifesta generalmente a causa di una esagerata produzione di composti tossici; al contrario, i fattori che ne determinano la produzione sono più o meno sempre gli stessi durante tutto l'arco dell'esistenza. Ciò che certamente si modifica in maniera sorprendente è, da un lato la incapacità di utilizzare i mezzi fisiologici per contrastare efficacemente la formazione dei radicali liberi, dall'altro la riduzione dell'attività dei sistemi deputati alla riparazione dell'organismo (situazione conosciuta come "stress ossidativo"). Più in particolare, durante il processo evolutivo sono andati sviluppandosi meccanismi intrinseci che, in condizioni normali, si oppongono con grande successo al costante danneggiamento cui siamo sottoposti a causa della formazione dei radicali liberi e che, nel loro complesso, vengono denominati sistemi e/o sostanze antiossidanti. Di questi fa parte anche la melatonina e, anzi, essa viene attualmente considerata la sostanza endogena dotata di maggiora attività protettiva nei confronti dello stress ossidativo. Questo perché la sua azione si svolge non è soltanto diretta contro i prodotti nocivi della ossidazione, ma anche indiretta, volta a stimolare l’attività degli altri componenti dei sistemi di difesa (ad esempio il glutatione) e ad inibire gli enzimi pro-ossidanti (come le lipossigenasi e l’ossido nitrico sintetasi)(Harderland, 1997). La capacità che la melatonina ha di interagire con i radicali liberi (fatto, questo, che le ha valso l'appellativo di "spazzino dei radicali liberi"), deriva sia dal fatto che tale ormone possiede una grande affinità chimica per questi composti, sia dalla capacità di reazione diretta anche nei confronti di altre sostanze nocive, quali gli anioni superossidi. In tal senso la melatonina agisce, contrariamente ad altri comuni anti ossidanti come il glutatione e l'acido ascorbico, in maniera definitiva sui radicali liberi, bloccando cioè la catena di eventi che conduce alla produzione e all'automantenimento delle sostanze ossidanti. In particolare, a livello cerebrale, è anche in grado di determinare la stimolazione dell'attività del più importante sistema antiossidante in quella sede e cioè l'enzima glutatione perossidasi. La protezione fornita dalla melatonina sembra essere diretta a livello della struttura maggiormente vulnerabile agli attacchi delle sostanze tossiche e cioè il DNA, legandosi ad esso, all'interno del nucleo delle cellule. A conferma di tale attività, nostri dati del tutto recenti indicano che la melatonina si localizza sia a livello del citoplasma che del nucleo delle cellule che costituiscono gli organi dell'apparato riproduttivo dell'animale maschio (Nordio et al., 2008 in press). D'altronde è stato anche documentato che la melatonina è in grado di proteggere i linfociti umani dai danni diretti al codice genetico determinati da insulti diversi, quali radiazioni, sostanze tossiche, radicali liberi, campi elettromagnetici, ecc.
Qualora la protezione fornita dalla melatonina si riduca, sia gradualmente a causa di una fisiologica diminuzione dei livelli di melatonina che si manifesta ad esempio con il progredire dell'età, sia rapidamente, in corso di stress cronici e di malattie debilitanti che determinano un impoverimento generale del potenziale antiossidativo dell'organismo, i soggetti risulteranno maggiormente esposti all'azione dei radicali liberi, particolarmente dannosi a livello cerebrale. In tale ottica, tutta una serie di lavori indicanti il ruolo protettivo svolto dalla melatonina nel corso di patologie degenerative cerebrali quali l’Alzheimer e il Parkinson. Nel primo caso, malgrado l’origine del processo neurodegenerativo sia ancora ignota, il danno determinato dall’aumento dei radicali liberi è stato identificato come uno dei processi patogenetici più potenti che conducono a morte le cellule neuronali. Tale particolare vulnerabilità del tessuto cerebrale dipende, almeno in parte, dal fatto che esso è ricco in fosfolipidi e proteine che sono molto sensibili al danno ossidativo e hanno un sistema antiossidativo di difesa scarsamente potente (Srinivasan, 2002). Come conseguenza di ciò, la somministrazione di melatonina è in grado di prevenire i danni del sistema nervoso centrale e del fegato dipendenti dall’avanzare dell’età (Tresguerres et al., 2008). Inoltre, la terapia con melatonina in pazienti con morbo di Alzheimer si è rivelata in grado di migliorare la qualità del sonno in questi pazienti, oltre a ridurre le anormalità degli altri ritmi circadiani e, in generale, di rallentare la progressione della malattia (Asayama et al., 2003), come verrà meglio specificato in un capitolo successivo. Tale miglioramento sembra essere tanto più evidente quanto più precocemente viene iniziata la terapia. Infatti, dati della letteratura indicano che, nei pazienti con Alzheimer di stadio molto avanzato, i recettori per la melatonina MT1 risultano estremamente ridotti. Pertanto, gli effetti della somministrazione di melatonina potrebbero essere del tutto inadeguati (Wu e Swaab, 2007). Per quanti riguarda il morbo di Parkinson, i dati della letteratura indicano ugualmente nel danno da ossidazione la causa della distruzione delle cellule cerebrali dopaminergiche. Anche i questo caso, la somministrazione di melatonina si è rivelata utile per i pazienti con Parkinson, aumentando la loro capacità di movimento e la durata del sonno (Dowling et al., 2005). Inoltre, dati recenti indicano che la somministrazione di melatonina in questi pazienti è in grado di migliorare la qualità del sonno (Mendes-Medeiros et al., 2007).
Melatonina e attività oncostatica
Ormai da diversi anni esiste evidenza che l’inizio e/o la progressione di alcuni tumori possano essere bloccati o quantomeno rallentati dall’incremento fisiologico notturno dei livelli di melatonina nel sangue e nel compartimento extracellulare (Blask et al., 2005, a). Inoltre, altri autori hanno potuto documentare che la somministrazione di melatonina è in grado di svolgere attività oncostatica in un’ampia serie di cellule tumorali, quali le cellule del carcinoma ovario, del carcinoma endometriale, del melanoma umano, del tumore prostatico e di quello intestinale (Pandi-Perumal et al., 2006). Per queste ragioni esiste consenso quasi generale nel ritenere che la melatonina abbia effettivamente attività oncostatica che viene svolta a più livelli. In particolare nel tumore mammario, la melatonina riduce la sintesi locale degli estrogeni, essenzialmente attraverso due meccanismi. Il primo che si esplica mediante una inibizione dell’asse ipotalamo-ipofisi-gonadi, con conseguente riduzione del livello degli estrogeni circolanti. Il secondo attraverso un’azione diretta sulle cellule tumorali, modulando sia l’attività trascrizionale dei recettori per gli estrogeni, sia le vie di comunicazione post-recettoriali, inibendo anche gli enzimi armatasi (controllano la conversione da androgeni ad estrogeni)(Kiefer et al., 2002; Dopfel et al., 2007). Tale ipotesi trova conferma, almeno per quanto riguarda il tumore della mammella, anche nell'uomo, infatti, in pazienti con neoplasia mammaria, sembra esistere una correlazione inversa fra contenuto di recettori per gli estrogeni delle cellule tumorali e picco notturno della melatonina. Più in particolare, un nostro studio di qualche anno fa riguardante pazienti di sesso femminile con patologia benigna e maligna della mammella, ha dimostrato che l'attività funzionale della ghiandola pineale risultava ridotta soltanto in presenza di cancro della mammella in stadio avanzato, indicando ancora una volta che la riduzione delle concentrazioni di melatonina in pazienti con tumore può interferire con la prognosi dei soggetti stessi (dati non pubblicati). Ulteriori dati indicano che l’attività oncostatica della melatonina si svolge anche tramite un effetto inibitorio sulla captazione degli acidi grassi (dell’acido linoleico in particolare) della cellula tumorale, allo scopo di prevenire la formazione di un composto di derivazione altamente mitogeno (l’acido 13-idrossioctadecadienoico), con la mediazione dei recettori MT1 ed MT2 (Blask et al., 2005, a). Le azioni anti tumorali della melatonina si svolgono anche tramite la sfruttamento del suo potenziale antiossidante in quanto essa è in grado di ridurre il numero delle mutazioni cellulari, stimolate dallo stress ossidativo (Karbownik et al., 2001). Inoltre, la melatonina, tramite la sua azione sul sistema immunitario attivando le citochine, inibisce la crescita tumorale. Infine, essa si è rivelata in grado di indurre apoptosi e di svolgere probabilmente un’azione anti-angiogenetica (Vijayalaxmi et al., 2002). Per quanto riguarda l’impiego della melatonina in associazione con terapie antiblastiche, essa è risultata in grado di aumentare la sopravvivenza e la qualità della vita in pazienti con carcinoma del polmone metastatizzato (Lissoni et al., 2003), riducendo maggiormente la progressione del tumore, rispetto all’uso della sola chemioterapia (Lissoni, 2007). Inoltre, l’associazione con altre terapie antitumorali si è dimostrata efficace, non soltanto nei confronti della progressione del tumore, ma anche nei confronti di un miglioramento della qualità della vita dei pazienti, migliorando la mielotossicità e linfocitopenia caratteristiche dei protocolli chemioterapici (Vijayalaxmi et al., 2002).
Altro aspetto importante nella valutazione del collegamento fra melatonina e tumori risiede nel fatto che i pazienti con vari tipi di carcinomi presentano un alterato ritmo di secrezione della melatonina (Grin e Grunberger, 1998). A conferma di ciò, un ampio studio molto recente che ha valutato il rischio dei lavoratori turnisti (infermiere) di sviluppare carcinoma della mammella, ha dimostrato che il lavoro notturno (attraverso l’esposizione alla luce durante la notte, associata ad una riduzione dei livelli di melatonina) aumenta il rischio di sviluppare carcinoma della mammella e del colon-retto (Schernhammer et al., 2006, a). Nello stesso studio, ai soggetti con livelli di melatonina più elevati era associato un rischio inferiore di sviluppare carcinoma della mammella. Questi dati così importanti vengono rafforzati da risultati ottenuti precedentemente nell’animale, indicanti che sangue di donne sane in premenopausa, prelevato durante le ore notturne e quindi contenente elevate quantità di melatonina, ha determinato l’inibizione della crescita di tumori indotti sperimentalmente. Allo stesso modo, il prelievo del sangue durante le ore diurne e la successiva somministrazione all’animale da esperimento determinava un incremento della velocità di crescita del tumore mammario (Blask et al., 2005, b).
Melatonina e sistema immunitario
La convinzione che la melatonina fosse in grado di interferire con il sistema immunitario risale a diversi anni or sono, quando uno studio ormai “storico” dimostrò che la inibizione della secrezione della melatonina determinava un’attenuazione della risposta immunitaria nell’animale da esperimento (Maestroni et al., 1986). Da allora ha preso il via tutta una serie di esperimenti per capire meglio in che modo la melatonina potesse essere collegata al funzionamento del sistema immunitario e attualmente è possibile affermare che le attività di immunostimolazione svolte dalla melatonina, a più livelli, sono state confermate in una grande varietà di organismi viventi, compreso l’uomo. In particolare, la inibizione della secrezione di melatonina, ottenuta mediante esposizione alla luce durante le ore notturne, oppure la eliminazione chirurgica della ghiandola pineale, determina una immunodepressione che coinvolge essenzialmente la capacità di produrre anticorpi in risposta ad antigeni cosiddetti T-dipendenti. Al contrario, la successiva somministrazione di melatonina all'inizio del periodo notturno si è rivelata in grado di ripristinare la qualità della risposta anticorpale, la resistenza all'attacco dei virus, dei parassiti e delle cellule tumorali, ed il peso del timo (Carrillo-Vico et al., 2005). In tal senso, la presenza di siti di legame specifici per la melatonina sulla superficie delle cellule linfocitarie indica un effetto diretto di tale composto sulla regolazione del sistema immunitario. Infatti, i cosiddetti linfociti-T, con la loro produzione di citochine, sembrano costituire, comunque, il principale mediatore degli effetti immunostimolanti della melatonina. Più in particolare, essa è in grado di attivare la produzione di interleuchine-2 e -4 da parte dei linfociti-T, la cui azione si svolgerebbe sulle cellule capaci di produrre gli elementi del sangue, fornendo loro la capacità di contrastare i danni provocati da una precedente esposizione a sostanze tossiche. Inoltre, la melatonina ha dimostrato di avere proprietà stimolanti la produzione di interleuchine anche da parte dei macrofagi (IL-2 e IL-12), oltre alla ben nota azione di stimolo dei linfociti-T. Inoltre, la correlazione dimostrata fra concentrazioni di melatonina e interleuchine-12 in pazienti HIV-positivi ha permesso di dimostrare che la riduzione della melatonina nel sangue di tali pazienti può contribuire all’alterazione della risposta immunitaria, soprattutto per quanto riguarda i linfociti T-helper1, il cui ritmo notturno è correlato con quello della melatonina (Nunnari et al., 2003; Carrillo-Vico et al., 2004). Tuttavia, gli effetti immuno-stimolanti della melatonina non dipendono soltanto dalla sua capacità di indurre la produzione di citochine, ma anche dalla capacità di stimolare la risposta immunitaria aspecifica (aumento del numero delle cellule “Natural-killer”: NK e dei monociti nel midollo osseo) svolgere azioni antiossidanti, antiapoptotiche e chemiotattiche (nei confronti delle cellule “T” e “B”)(Peña et al., 2007). Altro dato importante riguarda il fatto che le cellule del sistema immunitario sono una fonte non indifferente di melatonina, essendo state documentate elevate concentrazioni e la presenza degli enzimi per la sintesi di melatonina nel midollo osseo e nelle cellule immunocompetenti, anche dell’uomo (Carrillo-Vico et al., 2006).
Sulla base di tali considerazioni, alcuni autori suggeriscono che la melatonina possa svolgere un ruolo nella patogenesi di alcune malattie autoimmunitarie ed in particolare nell’artrite reumatoide i cui pazienti dimostrano livelli plasmatici di melatonina più elevati rispetto ai controlli. Gli stessi autori suggeriscono che l’incremento delle malattie autoimmunitarie che si verifica nei Paesi a latitudine elevata, durante il periodo invernale, potrebbe essere dipendente dall’aumento del quantitativo giornaliero di melatonina e dal conseguente stimolo dell’attività immunitaria (Maestroni et al., 2004).
FATTORI CHE INFLUENZANO I LIVELLI PLASMATICI DI MELATONINA
Esistono molteplici fattori in grado di modificare i livelli plasmatici di melatonina. Fra essi, la luce sembra essere il più efficace, ma anche temperatura, età, jet-lag, turnazioni, indice di massa corporea (maggiore la massa corporea, minore la secrezione urinaria di melatonina)(Schernhammer et al., 2006, b), abitudini alimentari, assunzione di farmaci possono modificare sia le caratteristiche del ritmo (avanzare o ritardare la fase del sonno), sia i livelli notturni di melatonina. Qui di seguito saranno riportate informazioni riguardanti tali fattori, esclusi quelli di cui si è già parlato in precedenza quali: la luce, il jet-lag e le turnazioni.
Melatonina ed invecchiamento
E' ormai generalmente accettato che il processo di invecchiamento sia associato alla degenerazione e alla desincronizzazione delle funzioni dell'organismo. Infatti, con l'avanzare dell'età, viene a ridursi drasticamente il potenziale di adattabilità alle variazioni dell'ambiente che ci circonda. In ultima analisi, e da un punto di vista puramente conservativo della specie, avendo superato la fase riproduttiva, l'organismo che invecchia viene messo nelle condizioni di diminuire fisiologicamente le proprie capacità di sopravvivenza e quindi dare spazio ad individui più giovani, in grado di riprodursi a loro volta, in modo da perpetuare la specie. E’ necessario sottolineare che, quando si parla di invecchiamento, non è possibile sottovalutare la grande differenza individuale fra i soggetti anziani in relazione, ad esempio, con il potenziale genetico dell'invecchiamento (cioè la predisposizione ad invecchiare più o meno velocemente, ereditata dai genitori), con la risposta ai fattori ambientali, con i livelli di melatonina, la cui maggiore o minore presenza può contribuire a spiegare il marcato divario che a volte si nota fra età cronologica ed età biologica del soggetto. Tale divario può essere evidenziato determinando quella che si definisce come la qualità dell'invecchiamento, per mezzo di una serie di accertamenti strumentali quali la valutazione del sistema cardio-vascolare, del fondo dell'occhio, della funzione respiratoria, le concentrazioni plasmatiche o urinarie di melatonina che ci daranno indicazioni precise sullo stato reale della capacità funzionale dei vari organi, indipendente dall'età anagrafica. In pratica, gli organi e di conseguenza le funzioni interne possono deteriorarsi meno rispetto a quanto non accada alla media delle persone della stessa età, consentendo all'individuo di apparire più giovane, sia di aspetto, sia come funzionalità endogena. Pertanto, dal momento che non invecchiamo tutti allo stesso modo, è verosimile pensare che la definizione di un solo fattore, sia esso genetico o umorale o altro, come causa essenziale del processo dell'invecchiamento non soltanto sia una spiegazione insufficiente, ma anche troppo semplicistica. In realtà, l'invecchiamento fisico e mentale sta diventando uno dei maggiori problemi di salute del nostro secolo. Ciò è dovuto al fatto che quasi sempre esso è accompagnato da una o più malattie concomitanti, definite croniche e/o degenerative le quali aggravano il normale cammino dell'invecchiamento. La loro comparsa è in genere grandemente favorita da abitudini di vita del tutto errate e protratte per un numero di anni sufficiente. A titolo di esempio, basti pensare alla cosiddetta “Sindrome metabolica”, all'arteriosclerosi, all'infarto, alla trombosi cerebrale, al cancro, ecc.: tutte patologie il cui momento iniziale può essere ricercato in una non più perfetta adattabilità dell'organismo (in termini di autodifesa) all'insulto provocato da alcuni stimoli nocivi e di solito perfettamente controllabili. Più in particolare, per quanto riguarda la sindrome metabolica, dati recentissimi ottenuti in una coorte di 40 pazienti, hanno dimostrato una correlazione fra alterazione dei livelli plasmatici di melatonina e genesi della malattia (Robeva et al., 2008). Da tali concetti ha preso vita un'ipotesi, definita teoria dell'invecchiamento, che tenta di spiegare che, almeno nelle prime fasi, invecchiamo essenzialmente perchè non siamo più in grado di eliminare con successo le sostanze tossiche derivanti, come già ricordato, dal processo di ossidazione e che momento per momento si formano nel nostro organismo o giungono dall'esterno e che si vanno, pertanto, accumulando progressivamente, innescando un lento ma inesorabile processo di degradazione.
In tutto ciò si inserisce la melatonina i cui livelli plasmatici notturni vanno via via diminuendo con il progredire dell’età, fino a più del 50% di riduzione rispetto ai livelli del giovane adulto. Un primo tentativo per spiegare come mai una struttura così potente riduca così drasticamente la propria capacità produttiva si basa sul rilievo, noto fin da tempi antichissimi, della comparsa di una progressiva calcificazione dell'epifisi, dovuta all'accumulo di sali di calcio all'interno dei pinealociti, denominata all’epoca "sabbia nel cervello". Attualmente, tale teoria non viene più ritenuta del tutto valida. Primo perchè un gran numero di soggetti anziani non presenta alcuna calcificazione della pineale; secondo, perché studi recenti ottenuti anche nel nostro laboratorio, hanno dimostrato che in pazienti psichiatrici di giovane età sono evidenziabili depositi di calcio all'interno della pineale; terzo, è stato dimostrato che la presenza di calcificazioni dei pinealociti non significa automaticamente riduzione della capacità di produrre melatonina (Bersani et al., 2002).
Pertanto sembra verosimile ipotizzare che la ridotta secrezione di melatonina propria dell'età avanzata dipenda, almeno in parte da una duplice serie di fattori: da un lato la riduzione intrinseca della capacità metabolica dei pinealociti a causa del progressivo logoramento di essi (fattori genetici, ambientali e di stile di vita); dall'altro, la concomitante alterazione di tutte le altre strutture cerebrali renderebbe queste non più in grado di fornire il numero sufficiente di segnali di cui l'epifisi ha bisogno per un corretto funzionamento.
Melatonina e mondo vegetale
La evidente ubiquità, unita al fatto che la melatonina è rilevabile in organismi così diversi tra loro quali ad esempio l’uomo e i batteri, suggerisce che essa sia parte integrante del processo evolutivo della natura fin dal suo inizio e quindi ne sottolinea l’importanza (Manchester et al., 1995; Maronde e Stehle, 2007). Inoltre, contrariamente a quanto ritenuto negli anni passati, è stato possibile documentare che la melatonina viene prodotta anche dalle strutture vegetali (Dubbels et al., 1995; Hattori et al., 1995) ed è quindi possibile variare il contenuto di melatonina dell’organismo mediante la loro introduzione con la dieta. Studi recenti indicano che essa non solo è presente anche negli organismi vegetali, ma risulta del tutto identica alla melatonina dell’animale, potendo raggiungere concentrazioni paragonabili a quelle plasmatiche, caratteristiche di un soggetto adulto. Pertanto è verosimile ipotizzare che la normale dieta sia la fonte di assunzione di quantità variabili di melatonina nell’individuo normale. Tralasciando il fatto che, ancor prima della nascita, l’individuo riceve informazioni sulle caratteristiche ritmiche dell’ambiente esterno attraverso la melatonina prodotta dalla madre (Okatani et al., 1998), è tramite il latte materno che si inizia ad acquisire melatonina dall’esterno (Illnerova et al., 1993). A questo punto è intuitivo pensare che anche il latte di altri mammiferi contenga melatonina e se ciò ha un significato relativo per quanto riguarda cavie o ratti, assume una valenza importante il considerare quello dei bovini e di altri animali che contribuiscono all’alimentazione umana. In effetti è stato possibile individuare la presenza di melatonina nel latte bovino e caprino, secondo il ben noto ritmo circadiano, con livelli più elevati (circa 30 pg/ml) in quello raccolto nelle prime ore del mattino (Eriksson et al., 1998). Dal momento che quest’ultimo è proprio quello che viene comunemente utilizzato dall’uomo a scopo alimentare e che la melatonina è una molecola alquanto stabile e resistente, si può dedurre che non soltanto il latte di per sè, ma anche alcuni dei suoi derivati contengano concentrazioni significative di melatonina. Discorso analogo può essere fatto anche per quanto riguarda la carne delle specie animali commestibili per l’uomo, anche se è necessario sottolineare che la cottura, specie se prolungata, distrugge la molecola dell’indolo.
Per quanto riguarda più strettamente il mondo vegetale, la melatonina (e anche il triptofano) è stata identificata ad esempio nel pomodoro, nel riso, nell’arancia, nella mela, nella banana, nelle ciliegie, nella buccia dell’uva nera, nel cetriolo, nelle alghe verdi, nelle mandorle ed anche in un gran numero di erbe medicinali. Le concentrazioni di melatonina sono risultate essere variabili, comprese fra i 2 e i 5300 pg/g (Dopfel et al., 2007). E’ interessante notare come, fra le erbe medicinali normalmente prescritte per ritardare gli effetti dell’invecchiamento oppure per contrastare i radicali liberi, ci siano tutte quelle a maggiore contenuto di melatonina (Panti-Perumal et al., 2006). Un accenno a parte merita il dato riguardante la identificazione della melatonina in un vegetale non comunemente impiegato nella dieta, ma con ben note proprietà terapeutiche: l’iperico (hypericum perforatum) o “erba di San Giovanni”. Da circa un millennio sono note le sue proprietà antidepressive e di miglioramento dell’umore, tanto che attualmente questo prodotto, venduto liberamente nelle erboristerie e farmacie, viene utilizzato per la terapia delle depressioni lievi e moderate. In particolare in Germania, dove circa il 30 % dei pazienti lo assume con successo al posto dei farmaci antidepressivi. Ciò che non era completamente chiaro riguardava il perchè del miglioramento qualitativo anche del sonno in questi pazienti. La recente documentazione di elevate quantità di melatonina in tale vegetale (circa 1700 ng/g) può essere la chiave di lettura per considerare che l’effetto dell’iperico sul sonno sia mediato dalla melatonina (Murch et al., 1997).
Riassumendo, quindi, è possibile sostenere che la melatonina, per le sue caratteristiche biochimiche, risulta presente in un gran numero di strutture e prodotti animali e vegetali, configurando in tal modo la concreta possibilità di una sua assunzione quotidiana con gli alimenti comunemente impiegati nella dieta. Pur se già ricordato, è tuttavia assai importante sottolineare ancora che la melatonina introdotta durante le ore diurne avrà una ridotta possibilità di svolgere le proprie azioni in quanto la sensibilità dei recettori per questa sostanza risulta minima in quel periodo. Al contrario, nel momento di massima sensibilità recettoriale e cioè verso sera, l’assunzione di alimenti contenenti melatonina può mimare il fisiologico ritmo circadiano endogeno, con la conseguenza di promuovere la ritmicità di processi fisiologici come il sonno (Dawson e van den Heuvel, 1998). Non a caso, è probabile che l’abitudine di assumere del latte la sera per favorire il sonno abbia, oltre ad una valenza psicologica certa, anche (o almeno in parte) la sua spiegazione scientifica nella presenza della melatonina che è in grado di indurre quelle modificazioni fisiologiche caratteristiche della fase di addormentamento (riduzione della temperatura corporea, sopore, ecc.).
Tali dati sulla diffusione della melatonina nel mondo animale e vegetale, associato a quelli considerati in precedenza e cioè la identificazione di siti di produzione alternativi all’epifisi (che resta comunque la sede con maggiore attività) e la facile diffusibilità all’interno delle cellule, sottolinea l’importanza di questo composto nel mantenere l’omeostasi degli organismi viventi, suggerendo nel contempo la necessità di abbandonare del tutto la definizione di ormone, ormai ristretta ed obsoleta, dal momento che la melatonina non soddisfa soltanto i criteri fondamentali per l’appartenenza al gruppo degli ormoni (sostanze di origine animale prodotte soltanto da cellule specifiche di una data struttura ghiandolare e liberate nel circolo periferico allo scopo di giungere ad una struttura-bersaglio lontana dal luogo di produzione).
Melatonina e farmaci
Dati internazionali dimostrano come, accanto a patologie da dipendenza quali l’etilismo ed il tabagismo, anche situazioni di inquinamento ambientale come l’esposizione a campi elettromagnetici (EMF) possano ridurre in maniera efficace il picco notturno di melatonina (Reiter, 1994; Burch et al., 1998). Inoltre, la considerazione che la somministrazione di farmaci comunemente usati in terapia possa determinare una riduzione (fino alla scomparsa) del picco notturno della melatonina, con tutte le conseguenze che conosciamo, apre un capitolo importante a conferma della frequente necessità di integrare i livelli plasmatici di melatonina. Una lunga serie di sostanze farmacologiche, infatti, è in grado di determinare una diminuzione delle concentrazioni plasmatiche notturne dell’indolo epifisario. In primo luogo i farmaci ?-bloccanti ed i FANS, ma anche i calcio-antagonisti, le benzodiazepine ed i corticosteroidi. L’assenza, o almeno la riduzione, del picco notturno di melatonina che essi determinano può causare la comparsa di disturbi del sonno. Il motivo di tale effetto è specifico per ciascun gruppo di sostanze: a) ?-bloccanti: blocco dei recettori ?-adrenergici sulla superficie del pinealocita e conseguente mancato stimolo alla produzione di melatonina (Brismar et al., 1988); b) FANS: blocco della produzione di prostaglandine, necessarie per il trasferimento del segnale extracellulare recettoriale verso l’interno del pinealocita (Murphy et al., 1994); c) calcio-antagonisti: blocco delle attività di trasferimento del segnale e di stimolo intracellulare operate dal corretto funzionamento dei canali del calcio (Meyer et al., 1986); d) benzodiazepine: anche una singola dose è in grado di sopprimere la secrezione notturna di melatonina (McIntyre et al., 1988); e) corticosteroidi: alcuni studi hanno dimostrato una correlazione inversa fra livelli plasmatici di tali sostanze e la melatonina, anche se non esiste universalità di vedute. Più in particolare, per quanto riguarda i corticosteroidi è necessario un ulteriore approfondimento in quanto il collegamento di essi con la melatonina si attua anche a livello del sistema immunitario (Vassiljev, 1994). E’ noto infatti che la melatonina svolga una potente azione immunostimolante, in grado di annullare l’effetto immunosoppressivo determinato dai corticosteroidi. Si intuisce quindi come una terapia con corticosteroidi possa avere un duplice ordine di effetti negativi in quanto, la riduzione della melatonina che ne deriva può determinare, da un lato la comparsa di alterazioni della qualità del sonno, dall’altro un’amplificazione dell’effetto immunosoppressivo dei corticosteroidi.
CENNI SULL’USO DELLA MELATONINA IN TERAPIA
Alla luce di quanto esposto in precedenza, è intuitivo il considerare la possibilità di integrare (o sostituire) la produzione di melatonina mediante la somministrazione esogena di tale molecola. Infatti, da più di un quarto di secolo la melatonina viene somministrata con successo a soggetti con disturbi del sonno e l’ambito di impiego terapeutico della melatonina si amplia progressivamente, man mano che vengono identificate nuove aree di attività dell’indolo pinealico. I primi risultati risalgono alla metà degli anni ’70 e da allora, un’enorme serie di dati ha permesso di<